I have been reading the report of an exploratory workshop from July 2021 on designing a biodiversity monitoring network across Canada to address priority monitoring gaps and engage Indigenous people across Canada. The 34 pages of their workshop report can be accessed here, and I recommend you might read it before reading my comments on the report:
https://www.nserc-crsng.gc.ca/Media-Media/NewsDetail-DetailNouvelles_eng.asp?ID=1310
I have a few comments on this report that are my opinion only. I think the Report on this workshop outlines a plan so grand and misguided that it could not be achieved in this century, even with a military budget. The report is a statement of wisdom put together with platitudes. Why is this and what are the details that I believe to be unachievable?
The major goal of the proposed network is to bring together everyone to improve biodiversity monitoring and address the highest priority gaps to support biodiversity conservation. I think most of the people of Canada would support these objectives, but what does it mean? Let us do a thought experiment. Suppose at this instant in time we knew the distribution and the exact abundance of every species in Canada. What would we know, what could we manage, what good would all these data be except as a list taking up terabytes of data? If we had these data for several years and the numbers or biomass were changing, what could we do? Is all well in our ecosystems or not? What are we trying to maximize when we have no idea of the mechanisms of change? Contrast these concerns about biodiversity with the energy and resources applied in medicine to the mortality of humans infected with Covid viruses in the last 3 years. A monumental effort to examine the mechanisms of infection and ways of preventing illness, with a clear goal and clear measures of progress toward that goal.
There is no difficulty in putting out “dream” reports, and biologists as well as physicists and astronomers, and social scientists have been doing this for years. But in my opinion this report is a dream too far and I give you a few reasons why.
First, we have no clear definition of biodiversity except that it includes everything living, so if we are going to monitor biodiversity what exactly should we do? For some of us monitoring caribou and wolves would be a sufficient program, or whales in the arctic, or plant species in peat bogs. So, to begin with we have to say what operationally we would define as the biodiversity we wish to monitor. We could put all our energy into a single group of species like birds and claim that these are the signal species to monitor for ecosystem integrity. Or should we consider only the COSEWIC list of Threatened or Endangered Species in Canada as our major monitoring concern? So, the first job of CAN BON must be to make a list of what the observation network is supposed to observe (Lindenmayer 2018). There is absolutely no agreement on that simple question within Canada now, and without it we cannot move forward to make an effective network.
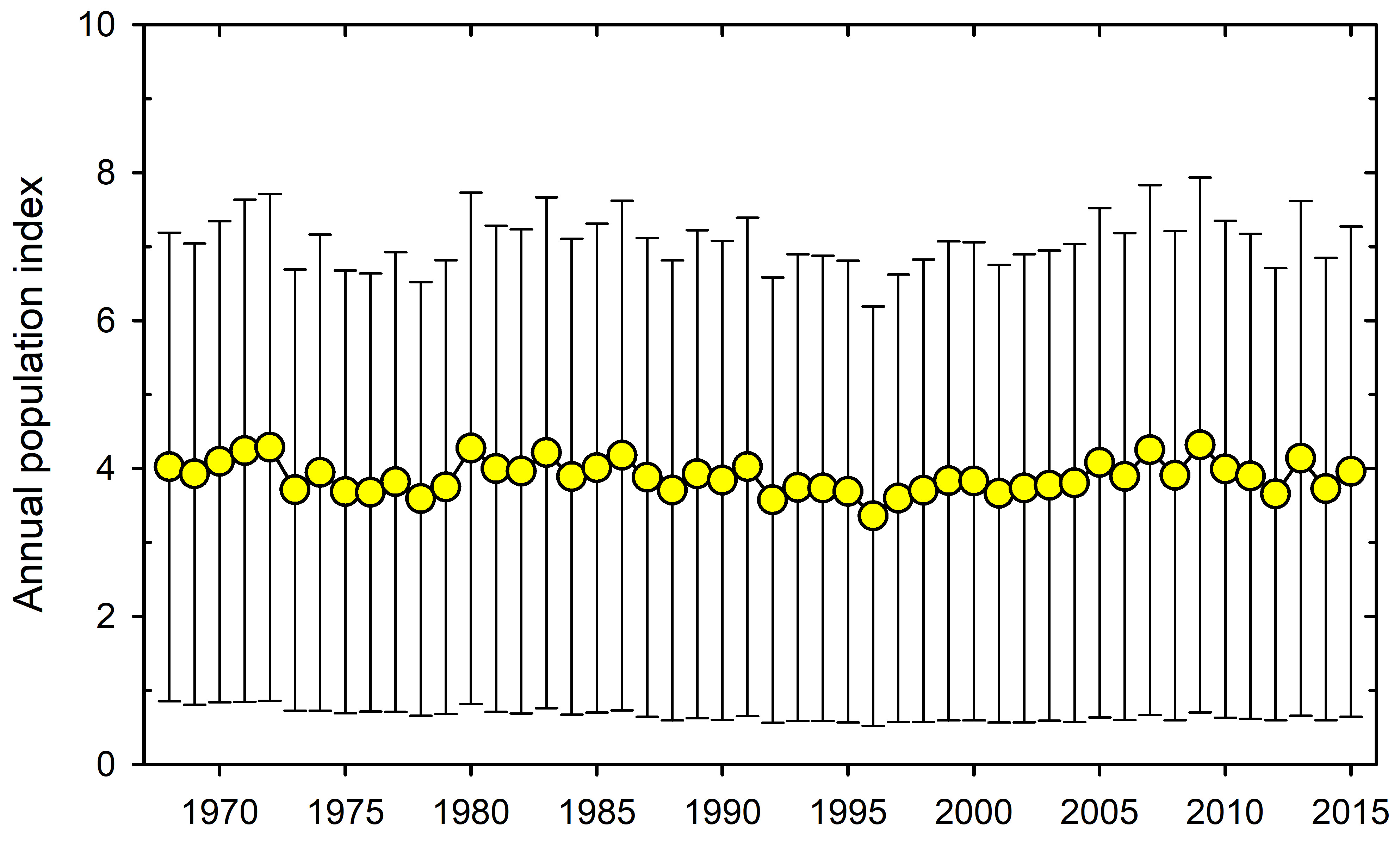
The second issue that I take with the existing report is that the emphasis is on observations, and then the question is what problems will be solved by observation alone. The advance of ecological science has been based on observation and experiment directed to specific questions either of ecological interest or of economic interest. In the Pacific salmon fishery for example the objective of observation is to predict escapement and thus allowable harvest quotas. Despite years of high-quality observations and experiments, we are still a long way from understanding the ecosystem dynamics that drive Pacific salmon reproduction and survival.
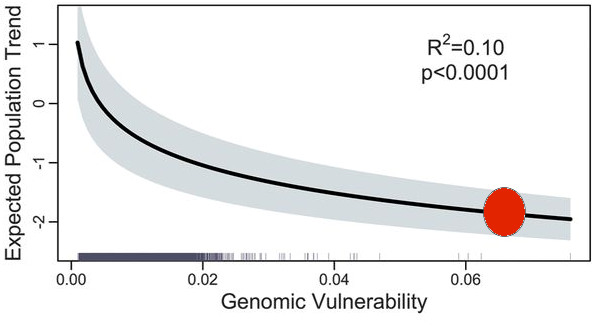
Contrast the salmon problem with the caribou problem. We have a reasonably good understanding of why caribou populations are declining or not, based on many studies of predator-prey dynamics, harvesting, and habitat management. At present the southern populations of caribou are disappearing because of a loss of habitat because of land use for forestry and mining, and the interacting nexus of factors is well understood. What we do not do as a society is put these ideas into practice for conservation; for example, forestry must have priority over land use for economic reasons and the caribou populations at risk suffer. Once ecological knowledge is well defined, it does not lead automatically to action that biodiversity scientists would like. Climate change is the elephant in the room for many of our ecological problems but it is simultaneously easy to blame and yet uneven in its effects.
The third problem is funding, and this overwhelms the objectives of the Network. Ecological funding in general in Canada is a disgrace, yet we achieve much with little money. If this ever changes it will require major public input and changed governmental objectives, neither is under our immediate control. One way to press this objective forward is to produce a list of the most serious biodiversity problems facing Canada now along with suggestions for their resolution. There is no simple way to develop this list. A by-product of the current funding system in Canada is the shelling out of peanuts in funding to a wide range of investigators whose main job becomes how to jockey for the limited funds by overpromising results. Coordination is rare partly because funding is low. So (for example) I can work only on the tree ecology of the boreal forest because I am not able to expand my studies to include the shrubs, the ground vegetation, the herbivores, and the insect pests, not to mention the moose and the caribou.
For these reasons and many more that could be addressed from the CAN BON report, I would suggest that to proceed further here is a plan:
- Make a list of the 10 or 15 most important questions for biodiversity science in Canada. This alone would be a major achievement.
- Establish subgroups organized around each of these questions who can then self-organize to discuss plans for observations and experiments designed to answer the question. Vague objectives are not sufficient. An established measure of progress is essential.
- Request a realistic budget and a time frame for achieving these goals from each group. Find out what the physicists, astronomers, and medical programs deem to be suitable budgets for achieving their goals.
- Organize a second CAN BON conference of a small number of scientists to discuss these specific proposals. Any subgroup can participate at this level, but some decisions must be made for the overall objectives of biodiversity conservation in Canada.
These general ideas are not particularly new (Likens 1989, Lindenmayer et al. 2018). They have evolved from the setting up of the LTER Program in the USA (Hobbie 2003), and they are standard operating procedures for astronomers who need to come together with big ideas asking for big money. None of this will be easy to achieve for biodiversity conservation because it requires the wisdom of Solomon and the determination of Vladimir Putin.
Hobbie, J.E., Carpenter, S.R., Grimm, N.B., Gosz, J.R., and Seastedt, T.R. (2003). The US Long Term Ecological Research Program. BioScience 53, 21-32. doi: 10.1016/j.oneear.2021.12.008
Likens, G. E. (Ed.) (1989). ‘Long-term Studies in Ecology: Approaches and Alternatives.’ (Springer Verlag: New York.) ISBN: 0387967435
Lindenmayer, D. (2018). Why is long-term ecological research and monitoring so hard to do? (And what can be done about it). Australian Zoologist 39, 576-580. doi: 10.7882/az.2017.018.
Lindenmayer, D.B., Likens, G.E., and Franklin, J.F. (2018). Earth Observation Networks (EONs): Finding the Right Balance. Trends in Ecology & Evolution 33, 1-3. doi: 10.1016/j.tree.2017.10.008.