Ecology progresses slowly when we have to study natural populations or communities. It is expensive to manipulate large units of habitat, and there are two approaches that suggest themselves to alleviate this problem. First, study small areas that can be analysed and manipulated by one or two persons. This can be a useful approach, depending on your question and hypotheses, and I do not discuss this approach here. The second approach is through experimental model systems. Typically this means taking the question or problem into a semi-laboratory system. For aquatic studies it may mean putting large cylinders in a lake (Carpenter 1996). For rodent studies it may mean putting populations into small fenced enclosures. For sake of clarity I will discuss this latter example with which I am familiar.
The key question for all experimental model systems in ecology is to know at what spatial and temporal scale the system works. To gain precision we typically want to conduct our studies within an enclosure of some small size. That is, we wish to study an open system with more precision by converting it to a closed system of some much smaller size. But what size allows the system to operate as an open natural population, in this example of rodents? In a sense we wish to know the shape of this generalized curve:
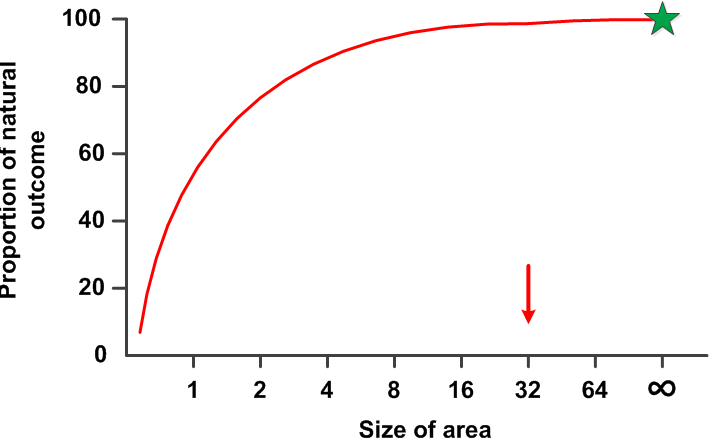
Assume there is some natural outcome known for the particular study. In the case of small rodents this might be that the population fluctuates in periodic ‘cycles’. The question then is what size of enclosure is needed to observe this same population trend. One simple way of looking at this is to ask for islands, what size of island allows a closed population to fluctuate in ‘cycles’. For this particular problem we know that you cannot observe ‘cycles’ in small rooms in the laboratory or even in 1 ha field enclosures.
Many other examples can be given for this type of question in ecology. For example, we may know that infanticide in a particular species is rare in natural populations. But if we raise the same species in small cages in the laboratory, we may observe infanticide very commonly. We would conclude that this is not the natural state of this system, and thus decide that you could not draw conclusions about the frequency of infanticide by studying it in small cages.
The critical judgement is whether any experimental model system we design will mimic natural processes that occur in open, real world populations or communities. All too often in ecological studies we assume that the size of the enclosure or study area that we are using is “natural” and the conclusions will represent what happens in natural populations or communities. In an ideal world we would examine a series of sizes of our study enclosures to see the best one that mimics natural outcomes. But this cannot always be done for reasons of time and money. In some cases we have no idea what the natural situation is, and in these cases it is most difficult to know if our model system results bear any relationship to reality.
This whole issue is another way of looking at the problem of habitat fragmentation – how small a piece of habitat can we get by with to conserve species X or community Y? These types of conservation questions always involve a temporal as well as a spatial dimension, given the problem of extinction debts (Krauss et al. 2010). In the extreme case we can argue that we can conserve at least some species in zoos, but this is a way of avoiding the main goal of conserving natural environments and processes.
The bottom line is to ask yourself as you are setting up a study using an experimental model system approach whether the process you are investigating can be observed at the spatial and temporal scale you have available. Alternatively it may be important to try to construct the curve shown above for the system of interest. This question is important because some previous studies for any ecological system may have reached invalid conclusions because of a faulty spatial scale of the model system.
Carpenter, S. R. 1996. Microcosm experiments have limited relevance for community and ecosystem ecology. Ecology 77:677-680.
Krauss, J., R. Bommarco, M. Guardiola, R. K. Heikkinen, A. Helm, M. Kuussaari, R. Lindborg, E. Öckinger, M. Pärtel, J. Pino, J. Pöyry, K. M. Raatikainen, A. Sang, C. Stefanescu, T. Teder, M. Zobel, and I. Steffan-Dewenter. 2010. Habitat fragmentation causes immediate and time-delayed biodiversity loss at different trophic levels. Ecology Letters 13:597-605.