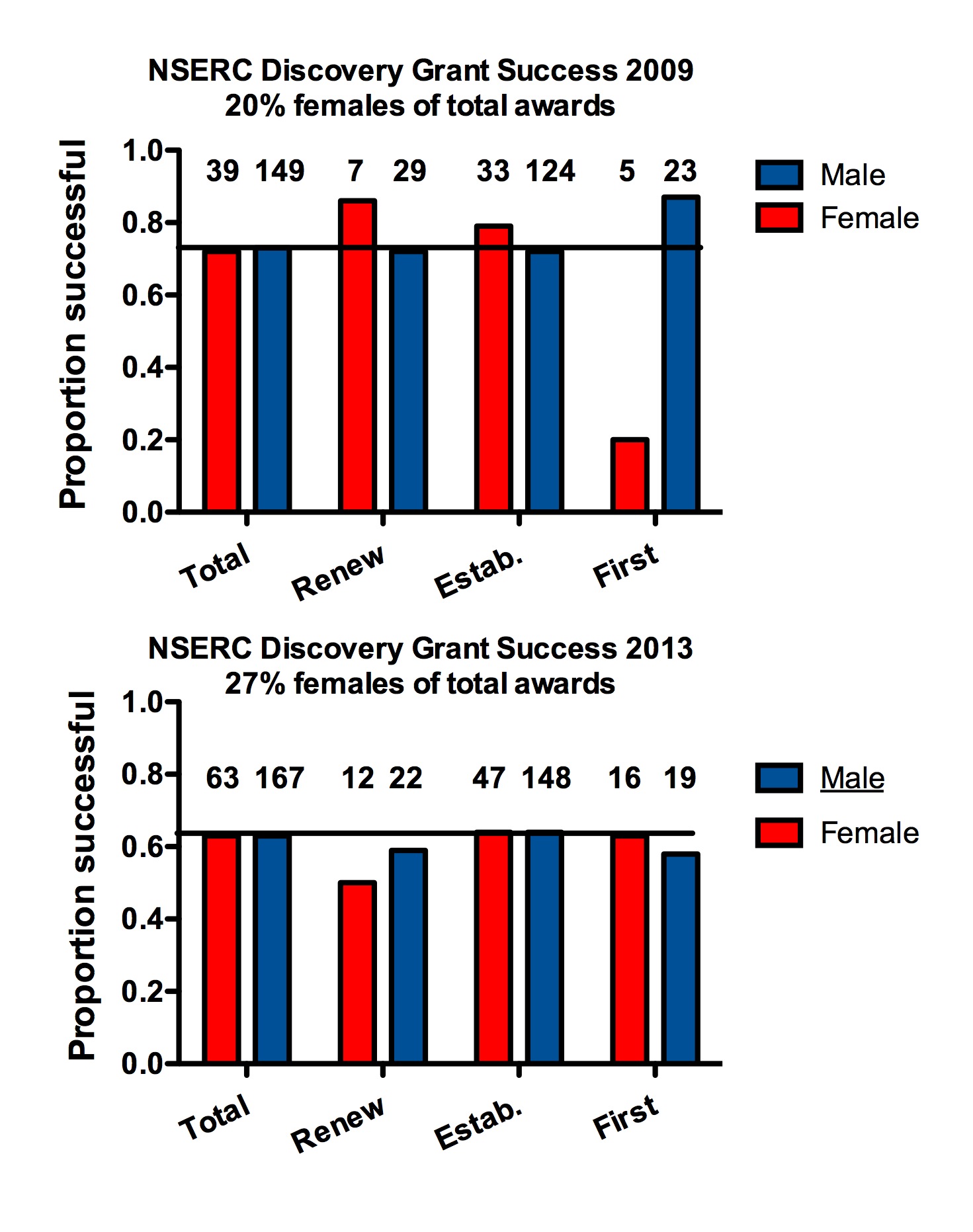
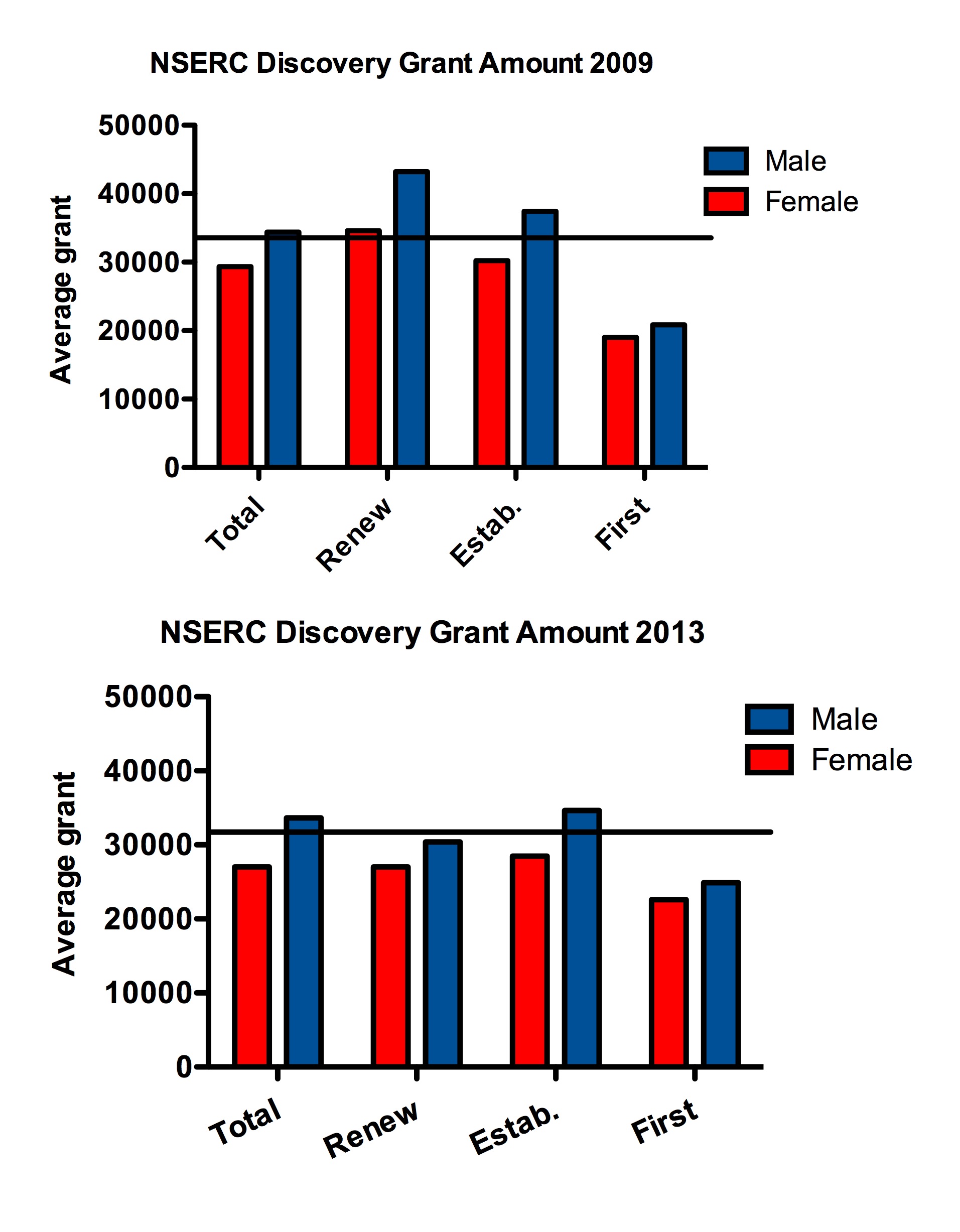
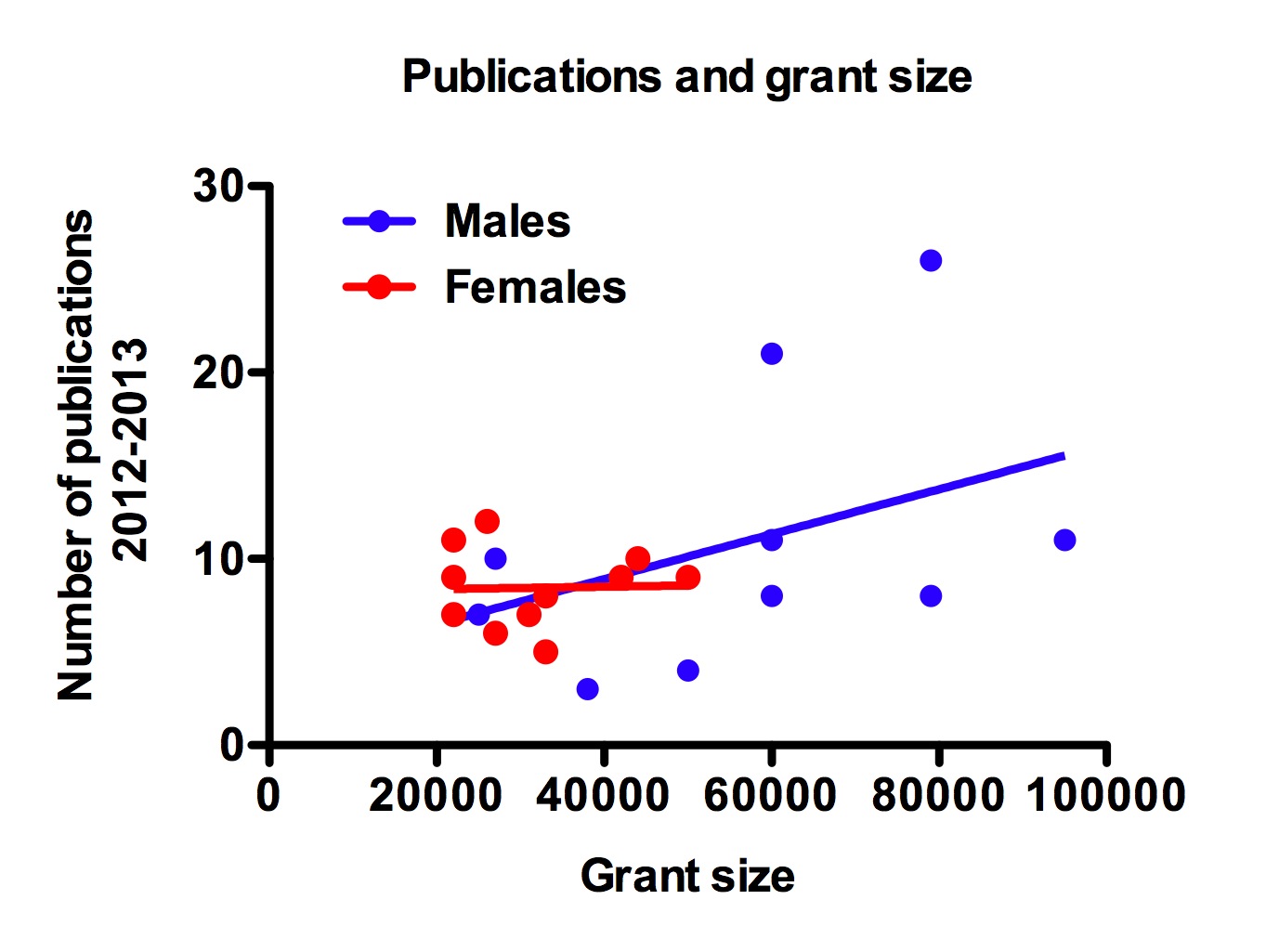
At the ESA meeting in 2014 Greg Dwyer (University of Chicago) gave a talk titled “Trying to understand ecological data without mechanistic models is a waste of time.” This theme has recently been reiterated on Dynamic Ecology Jeremy Fox, Brian McGill and Megan Duffy’s blog (25 January 2016 https://dynamicecology.wordpress.com/2016/01/25/trying-to-understand-ecological-data-without-mechanistic-models-is-a-waste-of-time/). Some immediate responses to this blog have been such things as “What is a mechanistic model?” “What about the use of inappropriate statistics to fit mechanistic models,” and “prediction vs. description from mechanistic models”. All of these are relevant and interesting issues in interpreting the value of mechanistic models.
The biggest fallacy however in this blog post or at least the title of the blog post is the implication that field ecological data are collected in a vacuum. Hypotheses are models, conceptual models, and it is only in the absence of hypotheses that trying to understand ecological data is a “waste of time”. Research proposals that fund field work demand testable hypotheses, and testing hypotheses advances science. Research using mechanistic models should also develop testable hypotheses, but mechanistic models are certainly are not the only route to hypothesis creation of testing.
Unfortunately, mechanistic models rarely identify how the robustness and generality of the model output could be tested from ecological data and often fail comprehensively to properly describe the many assumptions made in constructing the model. In fact, they are often presented as complete descriptions of the ecological relationships in question, and methods for model validation are not discussed. Sometimes modelling papers include blatantly unrealistic functions to simplify ecological processes, without exploring the sensitivity of results to the functions.
I can refer to my own area of research expertise, population cycles for an example here. It is not enough for example to have a pattern of ups and downs with a 10-year periodicity to claim that the model is an acceptable representation of cyclic population dynamics of for example a forest lepidopteran or snowshoe hares. There are many ways to get cyclic dynamics in modeled systems. Scientific progress and understanding can only be made if the outcome of conceptual, mechanistic or statistical models define the hypotheses that could be tested and the experiments that could be conducted to support the acceptance, rejection or modification of the model and thus to inform understanding of natural systems.
How helpful are mechanistic models – the gypsy moth story
Given the implication of Dwyer’s blog post (or at least blog post title) that mechanistic models are the only way to ecological understanding, it is useful to look at models of gypsy moth dynamics, one of Greg’s areas of modeling expertise, with the view toward evaluating whether model assumptions are compatible with real-world data Dwyer et al. 2004 (http://www.nature.com/nature/journal/v430/n6997/abs/nature02569.html)
Although there has been considerable excellent work on gypsy moth over the years, long-term population data are lacking. Population dynamics therefore are estimated by annual estimates of defoliation carried out by the US Forest Service in New England starting in 1924. These data show periods of non-cyclicity, two ten-year cycles (peaks in 1981 and 1991 that are used by Dwyer for comparison to modeled dynamics for a number of his mechanistic models) and harmonic 4-5 year cycles between 1943 and1979 and since the 1991 outbreak. Based on these data 10-year cycles are the exception not the rule for introduced populations of gypsy moth. Point 1. Many of the Dwyer mechanistic models were tested using the two outbreak periods and ignored over 20 years of subsequent defoliation data lacking 10-year cycles. Thus his results are limited in their generality.
As a further example a recent paper, Elderd et al. (2013) (http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3773759/) explored the relationship between alternating long and short cycles of gypsy moth in oak dominated forests by speculating that inducible tannins in oaks modifies the interactions between gypsy moth larvae and viral infection. Although previous field experiments (D’Amico et al. 1998) http://onlinelibrary.wiley.com/doi/10.1890/0012-9658(1998)079%5b1104:FDDNAW%5d2.0.CO%3b2/abstract concluded that gypsy moth defoliation does not affect tannin levels sufficiently to influence viral infection, Elderd et al. (2013) proposed that induced tannins in red oak foliage reduces variation in viral infection levels and promotes shorter cycles. In this study, an experiment was conducted using jasmonic acid sprays to induce oak foliage. Point 2 This mechanistic model is based on experiments using artificially induced tannins as a mimic of insect damage inducing plant defenses. However, earlier fieldwork showed that foliage damage does not influence virus transmission and thus does not support the relevance of this mechanism.
In this model Elderd et al. (2013) use a linear relationship for viral transmission (transmission of infection on baculovirus density) based on two data points and the 0 intercept. In past mechanistic models and in a number of other systems the relationship between viral transmission and host density is nonlinear (D’Amico et al. 2005, http://onlinelibrary.wiley.com/doi/10.1111/j.0307-6946.2005.00697.x/abstract;jsessionid=D93D281ACD3F94AA86185EFF95AC5119.f02t02?userIsAuthenticated=false&deniedAccessCustomisedMessage= Fenton et al. 2002, http://onlinelibrary.wiley.com/doi/10.1046/j.1365-2656.2002.00656.x/full). Point 3. Data are insufficient to accurately describe the viral transmission relationship used in the model.
Finally the Elderd et al. (2013) model considers two types of gypsy moth habitat – one composed of 43% oaks that are inducible and the other of 15% oaks and the remainder of the forest composition is in adjacent blocks of non-inducible pines. Data show that gypsy moth outbreaks are limited to areas with high frequencies of oaks. In mixed forests, pines are only fed on by later instars of moth larvae when oaks are defoliated. The pines would be interspersed amongst the oaks not in separate blocks as in the modeled population. Point 4. Patterns of forest composition in the models that are crucial to the result are unrealistic and this makes the interpretation of the results impossible.
Point 5 and conclusion. Because it can be very difficult to critically review someone else’s mechanistic model as model assumptions are often hidden in supplementary material and hard to interpret, and because relationships used in models are often arbitrarily chosen and not based on available data, it could be easy to conclude that “mechanistic models are misleading and a waste of time”. But of course that wouldn’t be productive. So my final point is that closer collaboration between modelers and data collectors would be the best way to ensure that the models are reasonable and accurate representations of the data. In this way understanding and realistic predictions would be advanced. Unfortunately the great push to publish high profile papers works against this collaboration and manuscripts of mechanistic models rarely include data savvy referees.
D’Amico, V., J. S. Elkinton, G. Dwyer, R. B. Willis, and M. E. Montgomery. 1998. Foliage damage does not affect within-season transmission of an insect virus. Ecology 79:1104-1110.
D’Amico, V. D., J. S. Elkinton, P. J.D., J. P. Buonaccorsi, and G. Dwyer. 2005. Pathogen clumping: an explanation for non-linear transmission of an insect virus. Ecological Entomology 30:383-390.
Dwyer, G., F. Dushoff, and S. H. Yee. 2004. The combined effects of pathogens and predators on insect outbreaks. Nature 430:341-345.
Elderd, B. D., B. J. Rehill, K. J. Haynes, and G. Dwyer. 2013. Induced plant defenses, host–pathogen interactions, and forest insect outbreaks. Proceedings of the National Academy of Sciences 110:14978-14983.
Fenton, A., J. P. Fairbairn, R. Norman, and P. J. Hudson. 2002. Parasite transmission: reconciling theory and reality. Journal of Animal Ecology 71:893-905.