![]()
Kin Selection &
Social Evolution
![]()
|
|
+ |
-- |
|
+ |
mutualism |
altruism |
|
-- |
selfishness |
spite |
Ü Mutualism + + : every individual benefits
Ü Selfishness + - : actor benefits, recipient pays cost
Ü Altruism - + : actor pays cost, recipient benefits
Ü Spite - - : every individual pays cost
![]() The paradox of altruism – why do something that reduces your own fitness, even if it helps another?
The paradox of altruism – why do something that reduces your own fitness, even if it helps another?
Darwin recognized as a challenge to his theory of evolution by natural selection.
Hamilton found the solution -- Help your kin.
Why is this a solution?
Take the gene-centered view: kin share genes, so helping kin also ‘helps’ an individual’s own genes. In other words, an allele that causes altruistic behavior will spread in a population if individuals help other individuals who carry that allele (because that increases the frequency of the allele).
Inclusive fitness. Individual fitness includes both the individual’s own reproduction (direct fitness) and increases in its relatives’ reproduction that are caused by the individual’s help (indirect fitness)
But how closely related do kin need to be?
And how much help should an individual provide?
![]() Hamilton’s rule:
Hamilton’s rule:
Two ways to think of it
r > c/b
relatedness should exceed the ratio of cost to benefit
c < rb
cost must be less than amount of benefit * relatedness
Generally, this means that altruism is likely to evolve when
Ex: Alarm calling in ground squirrels
Beldings ground squirrels live in colonies with burrows in close proximity. Females give calls to warn of approaching predators. This increases their risk of being caught by the predator, but decreases the risk for others in the colony.
So why risk this danger and call?
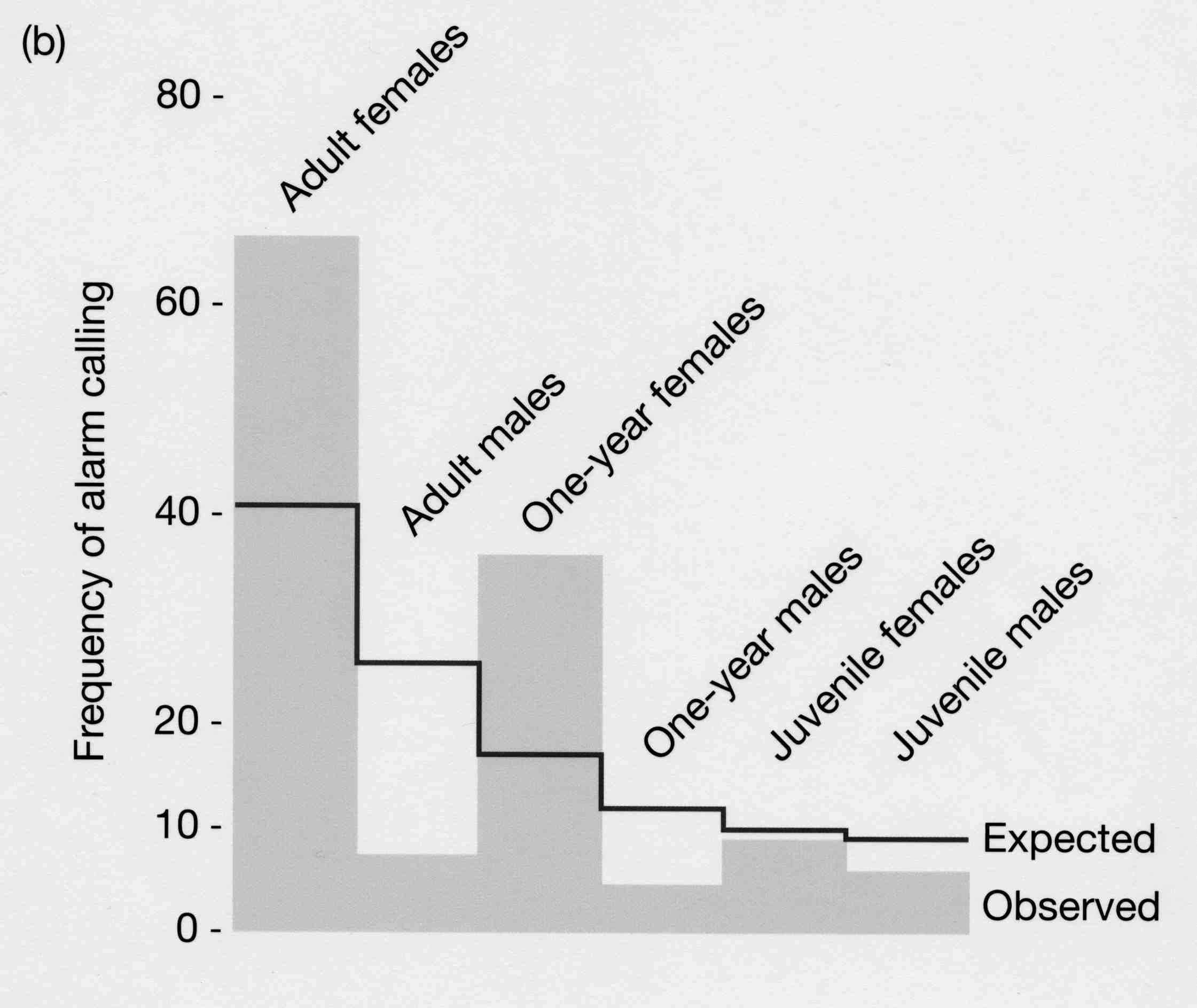
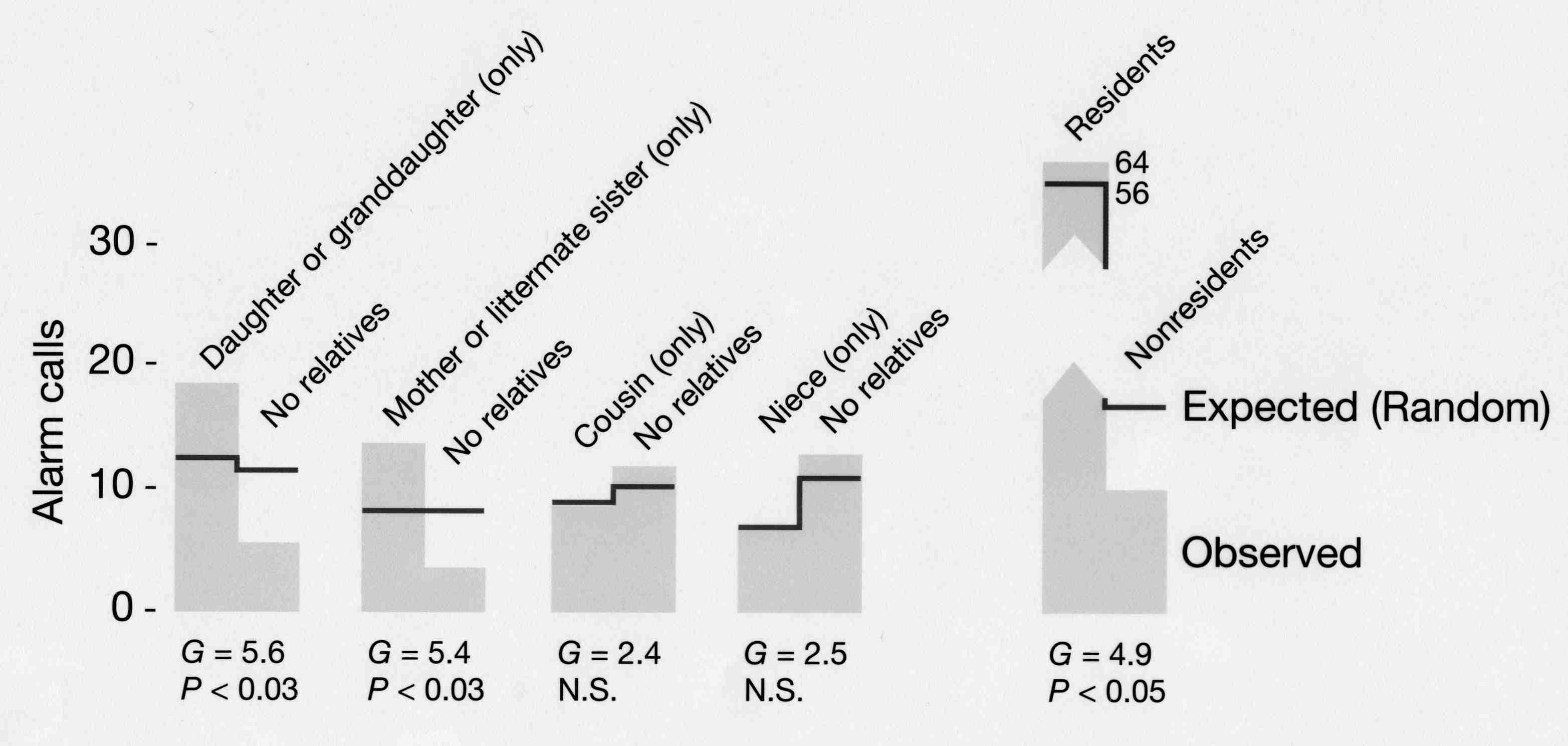
So, females may help relatives to avoid predation, thereby increasing the representation of their alleles in future generations. This means that alarm calling in the presence of relatives is favored.
Problem: recognizing kin
How to know if another individual is closely related?
Heuristic rules: Familiar individuals are more likely to be kin. This often works well, but not always.
Phenotype matching: Individuals with very similiar phenotypes are more likely to be kin. Traits have to be heritable and highly variable.
Signals to identify kin include vocal, chemical, visual, and morphological.
Calculating relatedness coefficients
Methods for estimating:
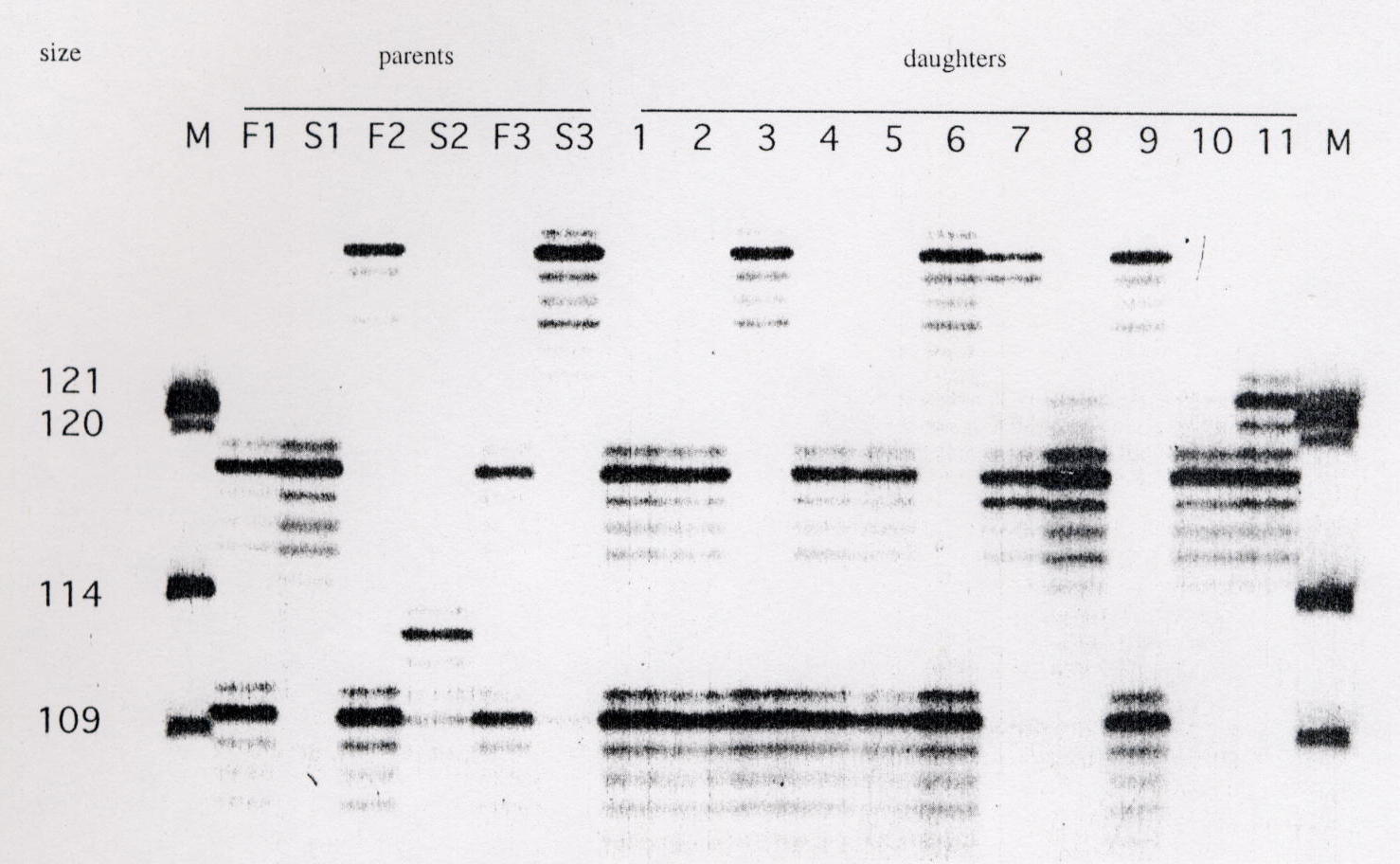
Assign maternity and paternity to offspring based on shared marker alleles.
Can also use for entire social groups or to compare relatedness among social groups.
Do not need prior information about relationships between individuals. Often useful in field studies and when relatedness estimates for many individuals is needed.
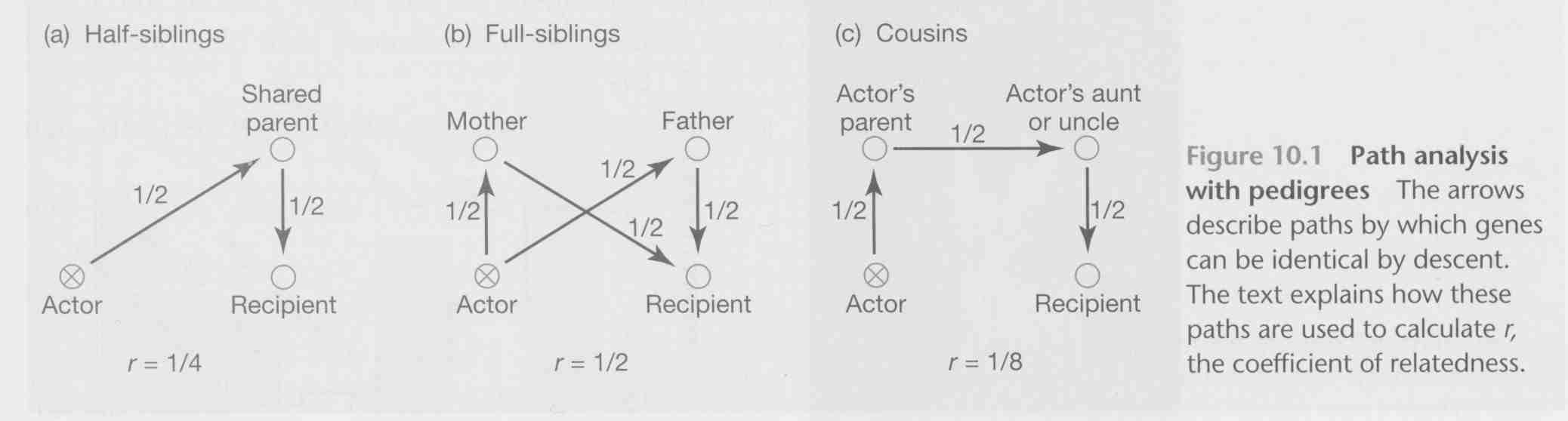
Calculate the expected proportion of shared alleles based on known relationships among individuals.
Trace paths of descent from actor to recipient, including all connections. The probability that shared parents transmitted the same alleles to both is found by multiplying each segment of the path connecting individuals. If actor & recipient share more than one parent, add values for each path to determine total probability of genes identical by descent (ibd).
Often used by breeders, or in long-term studies where individuals are marked. Cannot be applied in situations where pedigree is not known.
The evolution of eusociality
An extreme form of altruism -- Eusociality
Social organization with:
|
. |
|
Found in:
Social insects: bees, ants, wasps, termites
Naked mole rats
Workers completely forgo their own reproduction and help to raise their siblings. Their inclusive fitness consists entirely of indirect fitness. They have no direct fitness.
How could such an extreme form of cooperation evolve?
Haplodiploidy facilitates the evolution of eusociality because a female is more closely related to her sisters than to her own offspring. This means she increases her inclusive fitness most by helping to rear sisters.
Haplodiploidy: females are formed when eggs are fertilized and males are formed when eggs are unfertilized.
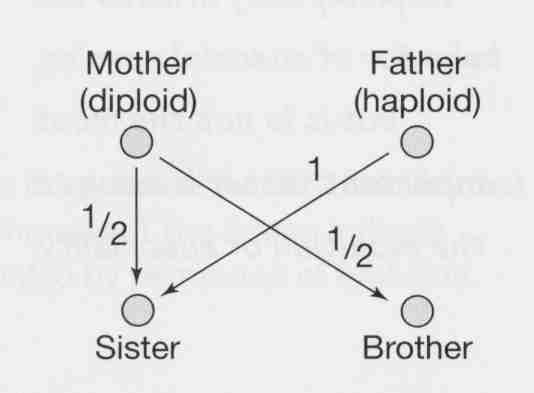
Relatedness coefficients
queen to female offspring 0.5
queen to male offspring 0.5
worker to worker 0.75
worker to own offspring 0.5
Ü But, not all haplodiploid taxa are eusocial, and not all eusocial taxa are haplodiploid.
This means that the evolution of eusociality depends on more than just relatedness.
Remember that Hamilton’s rule also has terms for benefits and costs, which are influenced by ecological factors. We need to consider the magnitude of benefits and costs & the role of ecology.
![]()
Social evolution
![]()
We can predict social organization based on ecological factors and Hamilton’s rule. Consider costs & benefits, relatedness.
Social structure shows wide variation:
We expect group living when there are benefits for survival or reproduction, which often depend on ecology. It’s especially likely among relatives.
![]()
Cooperation and conflict
![]()
Social life is not without conflict because the interests of individuals are rarely exactly equal. Helping oneself should be favored in many cases, because r = 1. Conflict of interests is apparent when we look closely at social groups.
Ex: Worker - queen conflict in social insects
In a normal diploid population the equilibrium sex ratio (# females to # males) is 1:1 (or 0.5). Why?
In haplodiploid eusocial insects, this no longer holds. Workers and queens have a conflict of interest over the best sex ratio (in reproductive offspring, not workers). What causes this conflict?
This leads to conflict of interest between queens and workers over the best sex ratio. Who wins?
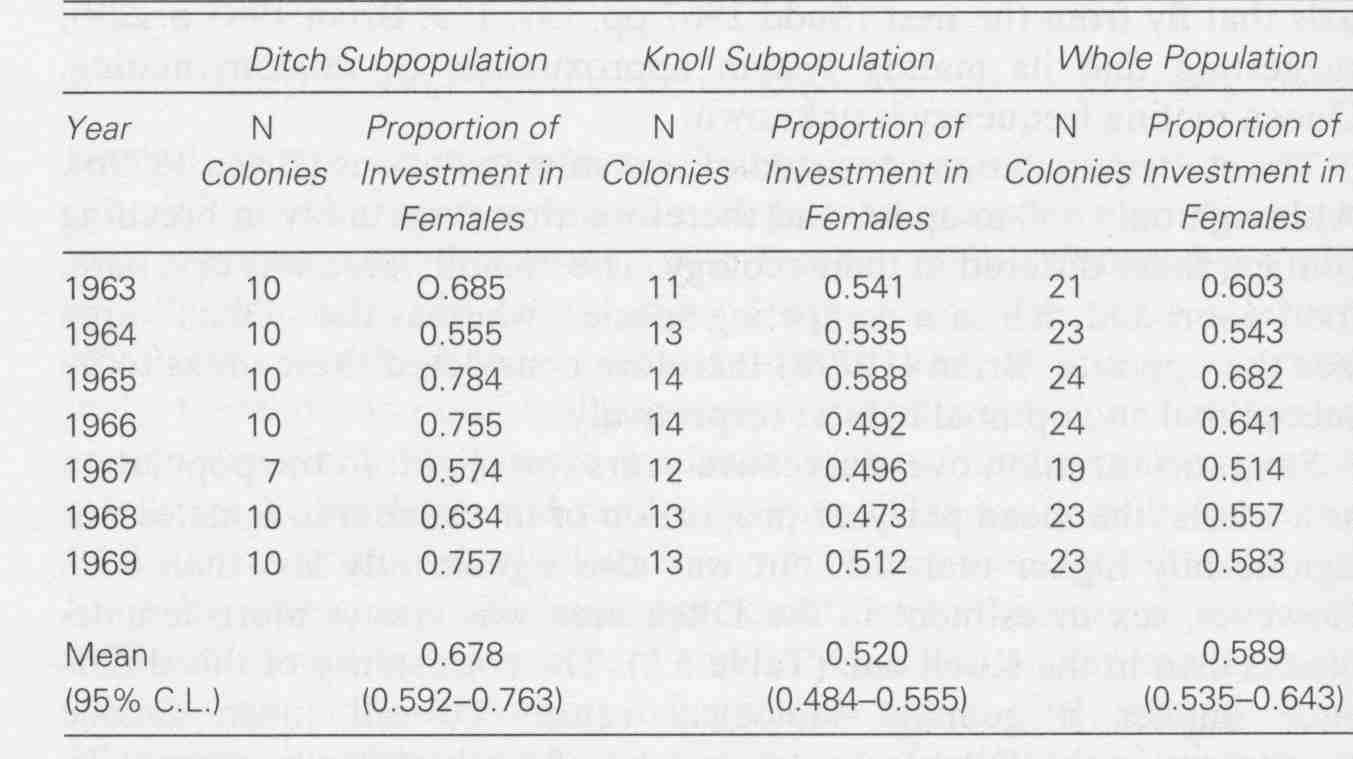
Two populations live in very different ecological conditions. Sex ratios differ.
Ex: Parent - offspring conflict
|
. |
|
The optimal period of parental care often differs for parents and offspring. Parents usually benefit by stopping care early. Offspring often benefit if parents continue care longer.
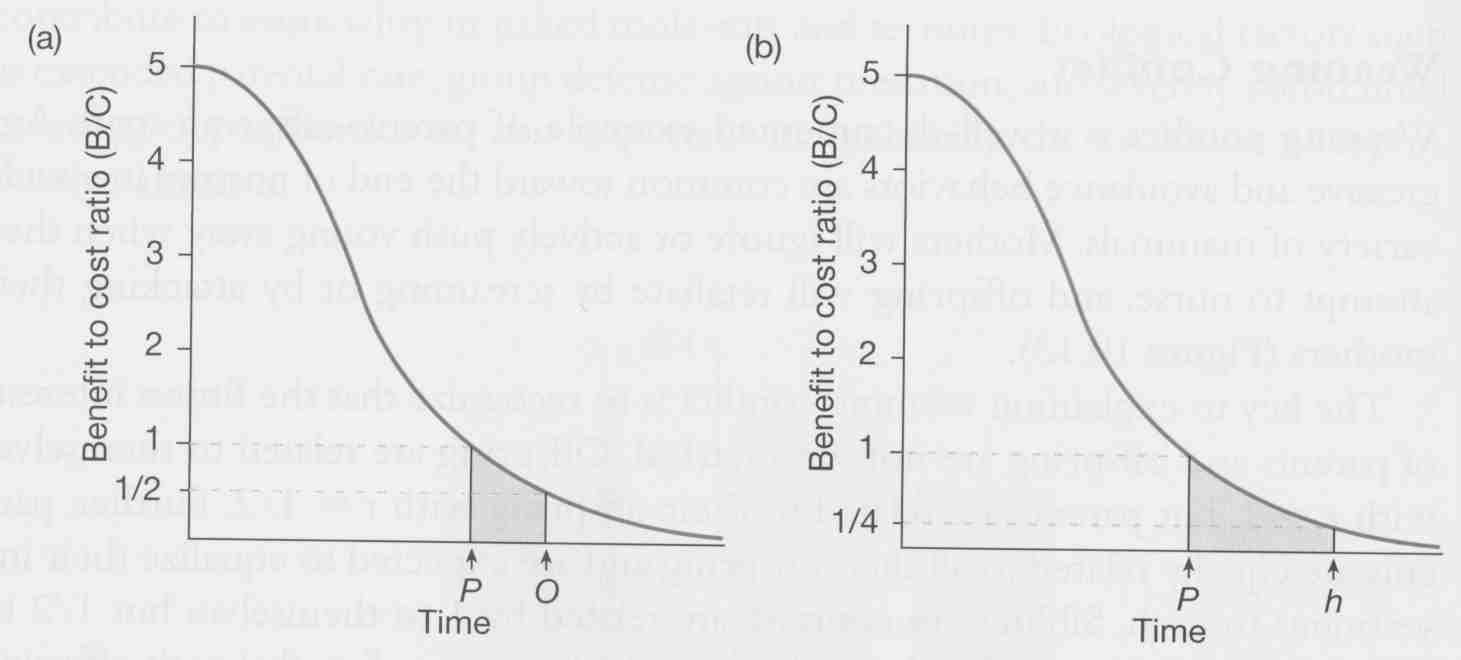
What leads to this conflict of interest?
What is optimum for parents & why?
Parents pay a cost for caring, and benefit by spreading their care over all their offspring, as long as the chance of survival stays high. Once offspring are able to survive on their own reasonably well, parents should stop providing care, saving their reproductive investment for future offspring. This should happen as soon as the cost/benefit ratio = 1.
What is optimum for offspring & why?
But offspring benefit by extracting as much care as possible, because it should always increase their survival by a little bit (and it doesn't cost them anything to receive the care).
The conflict arises because parents are equally related to all offspring by r = 0.5 and offspring are related to themselves by r = 1.0.
Ex: Evolution of cheating & cheater prevention
Cooperation is vulnerable to cheating, because individuals who cheat should have high fitness. They accrue benefits without paying the costs.
Prediction is that the frequency of cheaters should increase.
Ü Is cooperation evolutionarily stable?
BUT, with too many cheaters there will be no individuals to provide help and cheaters will do poorly (so will everyone else). Selection is frequency dependent. Cheating will stay at a low frequency.
AND, the ability to detect and exclude cheaters should evolve because cheaters lower fitness of cooperators.
Kin recognition is favored to detect cheaters. Individuals may be inspected to assess relatedness. Like a ‘password’ to allow entrance into a group.
EX: honeybee guards at the hive entrance
Also ability to recognize and remember individuals should be favored – remember who has helped.
Aggressively coerce potential cheaters to help, or punish for not helping.
EX: When they find a rich food source, rhesus monkeys give a call to alert others to the location of food. Individuals who do not call are attacked if they’re ‘found out’.
![]() These methods make cheating costly, so it no longer pays (in the evolutionary sense). This increases the cost to benefit ratio.
These methods make cheating costly, so it no longer pays (in the evolutionary sense). This increases the cost to benefit ratio.
Incorporating social structure into population genetics
|
|
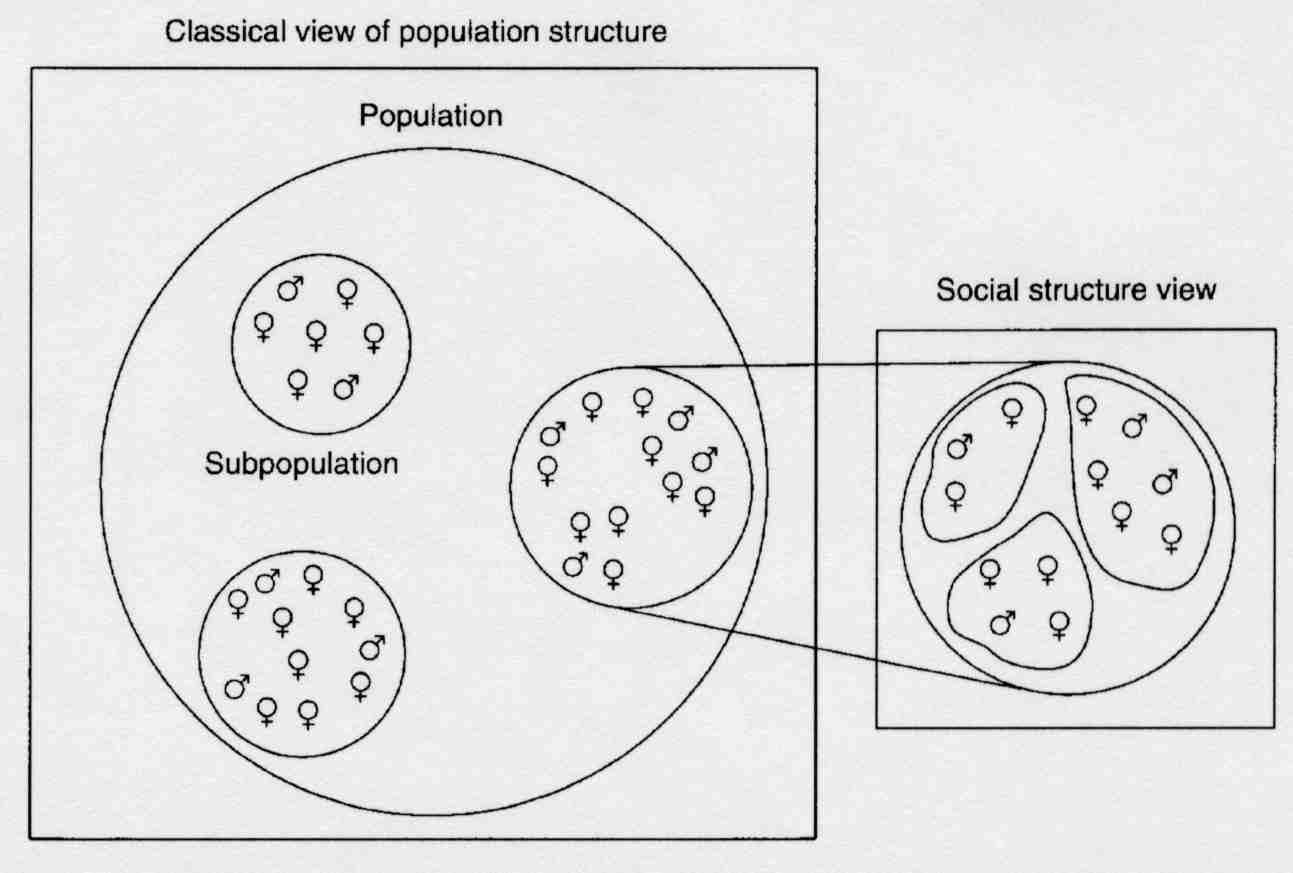
Social structure can result in genetic structure within populations
So far, models have assumed that individuals interact with others randomly. But in many species this is not the case. Social organization means that interactions are structured – behavior directed towards some individuals will be very different from behavior directed to others.
Not only do traits evolve that specifically facilitate social interactions, but the results of those interactions can change the evolutionary dynamics of other traits too.
EX: What happens if all mating is within groups?
Inbreeding is high so genetic variation within groups is very low, while genetic differences between groups is high.
The way genetic variation is distributed in populations is altered.