
The Origin of Species
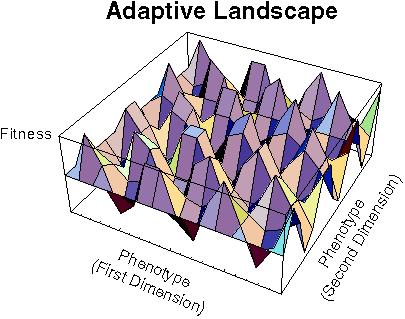
"...the living world is not a formless mass of randomly combining genes and traits, but a great array...of gene combinations, which are clustered on a large but finite number of adaptive peaks."Theodosius Dobzhansky (1951)
Why do living organisms cluster into discrete species?
What are the processes at work that lead to the origin of discrete species?
(The question that Darwin (1859) called "that mystery of mysteries".)
In this lecture, we will examine a number of different modes by which speciation may happen in nature.
These have been classified by Mayr (1963) as:
1B. Parapatric speciation (with contiguous but non-overlapping distributions)
1C. Sympatric speciation (with overlapping distributions)
2. Instantaneous speciation
2B. Cytological changes
3. Hybridization (producing new reproductively isolated species)
Our primary focus will be on how reproductive isolation can evolve under the gradual modes of speciation.
"Allopatric speciation is the evolution of genetic reproductive barriers between populations that are geographically separated by an extrinsic, physical barrier such as topography, water (or land) or unfavorable habitat."Futuyma (1998) p. 482
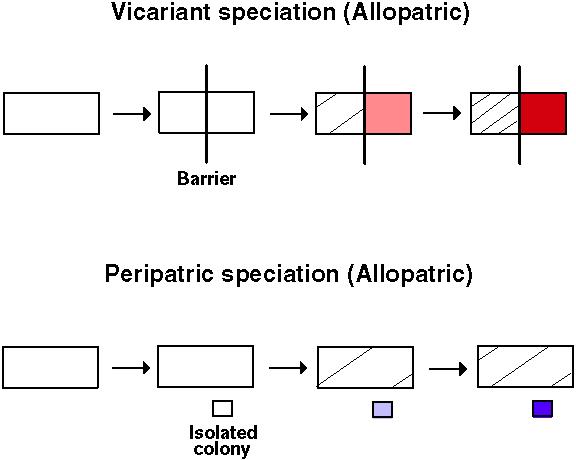
The extrinsic barrier halts gene flow between the isolated populations.
Without gene flow, each population follows an independent evolutionary trajectory, accumulating genetic changes through drift and/or selection.
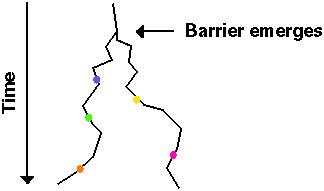
For these changes to lead to speciation, the genetic changes in one population must be incompatible with the genetic changes in the other population.
Allen Orr (1995) noted that as populations accumulate changes over time (R), the number of possible ways in which an incompatibility can occur rises at least as fast as R2: the more changes occur in one population, the more possible ways in which a genetic incompatibility could occur with the second population.
Once genetic incompatibilities have arisen between two separately evolving populations, the populations are no longer able to cross and produce fertile offspring.
 Speciation.
Speciation.
The genetic changes that accumulate in the two populations may be caused by natural selection, sexual selection, or by random genetic drift.
Which forces would you expect to be faster?
Which forces would you expect to be relatively more important in peripatric speciation?
Example: Isthmus of Panama
About 3 million years ago, North and South America merged, forming a land bridge (the Isthmus of Panama) and isolating the marine communities of the Carribean and Pacific.
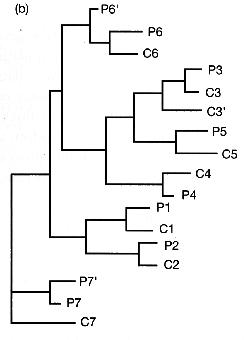
| Knowlton et al (1993) studied snapping shrimp from seven pairs of similar species on either side of the isthmus. |

|
|
A phylogeny of the seven pairs indicated that each of the species was most closely related to a similar looking species from the other coast. |
Mating experiments in the lab indicated that, in each case, the sister species from the Carribean and from the Pacific failed to produce viable offspring.
The formation of the land bridge between North and South America led to the isolation of populations on either side. With gene flow cut off, the populations diverged, leading to new species in 7/7 cases.
"Parapatric speciation is the evolution of reproductive isolation between populations that are continuously distributed in space, so that there is substantial movement of individuals, and hence gene flow, between them."Futuyma (1998) p. 498
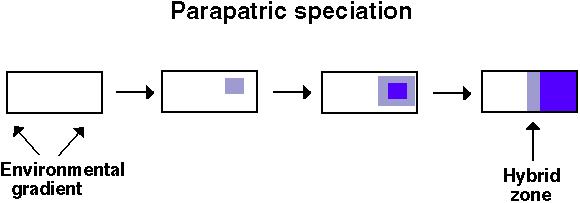
Without an extrinsic barrier to gene flow, genetic differences that arise within a population may be swamped by genetic exchange.
If, however, the population is distributed over space and:
If the habitat changes fairly abruptly, there may be a sharp border between different types: a hybrid zone.
Over time, the hybrid zone may shrink or remain stable.
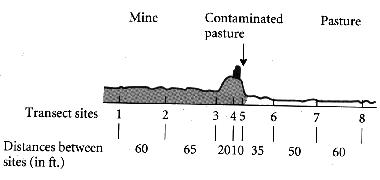
Example: Heavy metal tolerance
Antonovics and co-workers studied heavy metal tolerance in grasses growing near mines on land contaminated with lead and zinc.
| Plants from the grass species Anthoxanthum odoratum were tolerant near the mine but remained intolerant at distant sites. |
|
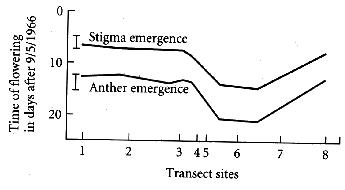
| Heavy metal tolerance was not the only difference observed; flowering time also differed along the transect. |
|
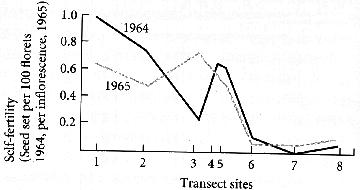
| Selfing rates were also higher near the mine. |
|
The divergence in flowering time and increased selfing rates have reduced gene flow between tolerant and non-tolerant grasses and increased the reproductive isolation of grasses near the mine.
In this case, heavy metal tolerance has evolved recently, during which time the species has always had a continuous distribution.
Therefore, parapatric speciation processes can be inferred. In most other cases, however, it is difficult to know whether a species that is currently parapatric may have been previously allopatric.
"Sympatric speciation is the evolution of reproductive isolation within a randomly mating population."Futuyma (1998) p. 498
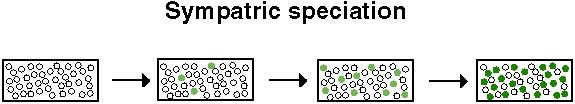
Sympatric speciation is the most controversial of all the modes of speciation, since divergence must occur in the face of high levels of genetic exchange.
Imagine a single species utilizing a resource that could support a wider variety of individuals.
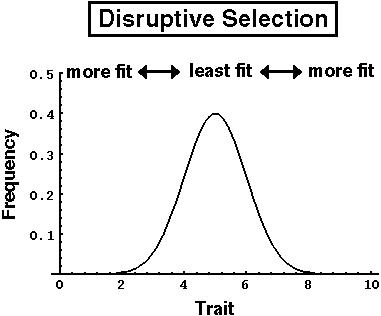
If the extreme phenotypes in a population have the highest fitness (disruptive selection), the population will not split apart if mating is random.
If, however, genes arise that cause extreme individuals to assortatively mate (= mate with similar types), then more extreme offspring can be produced:
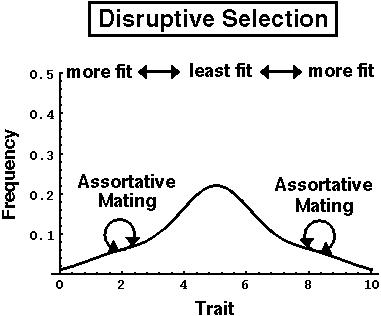
The problem with this idea is that it is difficult for assortative mating to evolve in a way that is coupled with the trait subject to disruptive selection.
One way for this coupling to be achieved is if there is tight linkage (low recombination) between the genes causing the trait and the genes causing assortative mating.
A second, more likely way is if the traits themselves lead to assortative mating.
For example, if the traits involve utilizing different resources and mating tends to happen around resources, then individuals with extreme traits will naturally mate with similar individuals.
Over time, if sufficiently strong assortative mating evolves, sympatric speciation will be the result.
Example: Apples and hawthorns
| The classic case of sympatric speciation is that of the apple maggot fly, Rhagoletis pomonella. |

|
Hawthorns were the original host plant of Rhagoletis pomonella until about 150 years ago, when the fly was observed on cultivated apple trees (introduced to the Americas).
The apple maggot fly is now widespread in the northeastern US and causes millions of dollars of damage each year to apple crops.
Rhagoletis courts, mates, and lays eggs on its preferred host plant.
Individuals that changed their preferred food source to apple trees (= a host shift), consequently tend to mate with other individuals who prefer apples.
Since the host shift, flies that prefer hawthorns and apples have diverged genetically.
Feder et al (1988, 1990) found statistically significant differences in allele frequencies at six loci, differences that allow the two populations to be identified.
Changes in the timing of mating have also occurred: mating on apples occurs ~ 3 weeks earlier.
Overall, matings between hawthorn and apple flies has been reduced to only ~6% and the two are now considered incipient species.
The exact mechanisms by which reproductive isolation is accomplished vary but fall into two main classes:
Premating isolation may be accomplished by: changes in habitat preferences, changes in timing of reproduction, physical incompatibilities, changes in mating preferences or mating behaviors.
2. Postmating (or zygotic) isolating mechanisms: Mating occurs but hybrid offspring are inviable, infertile, or produce inviable/infertile offspring.
Would you expect premating isolation to play more of a role in allopatric or sympatric speciation?
Coyne and Orr (1997) examined the causes of reproductive isolation in a number of sister species in Drosophila, finding:
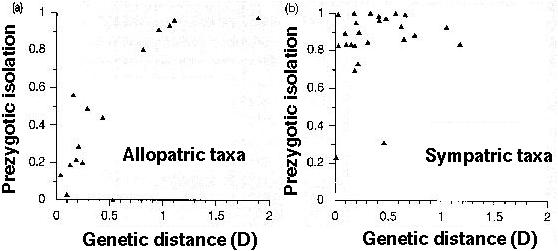
More rapid and complete prezygotic isolation between sympatric species pairs.
Would you expect premating or postmating isolation to evolve sooner between sympatric pairs?...between allopatric pairs?
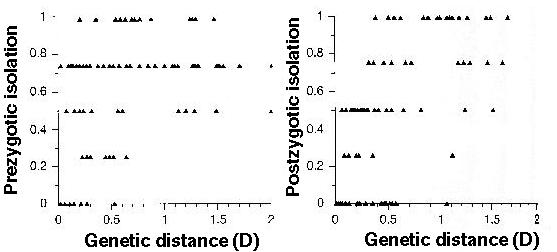
Prezygotic isolation evolves earlier in the speciation process, primarily due to the rapid mating isolation that evolves between sympatric species pairs.
Although examples of each mode of speciation are known, it remains unclear how frequently each occur.
Many evolutionary biologists suspect that allopatric speciation is responsible for the vast majority of speciation events.
SOURCES:
 Back to Biology 336 home page.
Back to Biology 336 home page.