Functional
Responses, Holling’s Disk Equation, and Predation Modeling Outline
Predators may respond to changes in prey-abundance in
several ways:
·
by changing individual behaviour in the short-term
(functional response)
·
by learning, up to the scale of a predator-lifetime
(developmental response)
·
by grouping at sites of high prey abundance
(aggregative response)
·
by reproducing to take advantage of a growing prey
population (numerical response)
Holling’s Disk Equation model focuses on the first
option: how does a predator respond to changes in prey density (assessed by
the number of prey which can be captured per foraging period at different
densities)?
Assumptions: (many
are unrealistic, but we’ll deal with this later)
w
the model looks at a single individual predator in
isolation
w
only one prey type (species) is being attacked
w
all prey individuals are identical/interchangeable (no
age/sex/health/etc. variation)
w
the predator thus takes the same period of
“handling-time” to deal with each prey
w
the number of prey caught is very small compared to
the number available (i.e. no depletion)
w
the predator is as hungry at the end as it was at the
start (hunts just as hard)
w
the total time available for foraging (per day, for
instance) is constant
The model
Call the density (individuals per unit area) of
prey available N . This is the main input-variable.
Call the total time spent foraging in a day T.
(a constant for a given forager, varies by species)
Call the total time spent searching for prey Ts ; this
will be made up of one or more variable-length bouts of searching, some ending
with prey-capture and some not.
Call time spent handling a single prey item Th . For a
given pair of species, this is a constant.
A forager may catch no prey, or some
prey, during a foraging period T; call the number of prey
caught (eaten) Ne . This
will be the main result-variable in the model. Thus the total time spent
handling (eating) prey will be Ne(Th) .
During the foraging period, the foraging predator
will be spending all of its time either searching for prey, or
handling captured prey. Thus the total foraging time is divided as follows:
![]() ( or, as we need below:
( or, as we need below: ![]() )
)
Now consider, just using common sense, what might
determine the number of prey caught (Ne):
n
the more time spent searching (larger Ts), the
more prey could be caught
n
the more prey available in the area (prey density, N),
the more prey could be caught
n
the more “behaviourally efficient” the predator, the
more prey could be caught
This last factor, efficiency, can be modeled
as a coefficient or multiplier: call it C .
So we could roughly model the number of prey
caught as:
![]()
We already derived above an expression describing Ts in terms
of our other variables, so:
![]() becomes
becomes ![]()
by direct substitution. If we multiply through to
remove the brackets:
![]()
and then rearrange to get both of the terms
containing Ne on the same side:
![]()
we can remove a common factor of Ne on the
left side of the equation:
![]()
and then “cross-multiply”, moving the bracketed item
over to the right side, thereby isolating the “result-variable” or dependent
variable, Ne, on the left:

and finally rearrange to get the N term
at the far right, since prey density is the model’s focus after all, and
we usually want to show the main independent variable as the last one in
the set of variables:
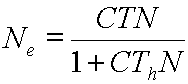
This is the basic form of the final disk-equation
model, in a “y = a x”-type format.
Using the model
Krebs shows a graph on page 222 (Figure 13.16), and
we will be drawing more graphs during lecture for further development of
the ideas. This is part of the model’s use.
Different mathematically-calculated forms of the
model (using the square of density, for example) can be used to generate
different curve-shapes. We will not be going in this direction in
Biology 303.
Another use of the model is (as with all models) to
“explore the extremes” – what happens when the model is applied to extreme
cases? Does it produce a biologically meaningful outcome? It is very
much worth our time to do this. Here is a start.
Consider the following cases:
What happens when N is very small?
Since
N measures density rather than absolute numbers, it can be a very
small value, easily much less than 1 (e.g. 0.005 individuals/km2). If this
is the case, then the CThN
term in the denominator becomes small compared to the “1” (so the denominator
approaches a value of simply 1, and can be ignored), and the whole equation
approaches:
![]()
This should make a lot of sense: if few prey are
present, almost all of T will be spent searching, but of course
most of that will be unsuccessful searching! At the extreme, a forager
might catch zero prey per time T, no matter its efficiency. Small
N thus gives very small Ne .
What happens when N is very large?
As
the N gets large, the “1” in the denominator becomes fairly insignificant
compared to the CThN
term, and so the model approximates:
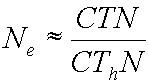 , which simplifies by canceling to:
, which simplifies by canceling to:
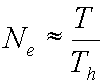
What does this mean? When the available prey are
abundant, hardly any time need be spent in searching for each one;
essentially the forager is surrounded by prey, which need only be picked up and
handled. Thus nearly the entire T period is composed of
handling-periods, and there will be a maximum number of prey which can be consumed,
since T and Th are both
considered constants – basically an asymptote. This can be interpreted as
evidence of saturation (but not physiological satiation, remember the
model’s assumptions don’t allow that!).
You might want to think of other ways of stretching
the model to extremes, as practice.
Limitations of the model
Basically all of the assumptions! The functional
response model would work only if all the assumptions outlined above were met,
and we know from basic commonsense principles that in many cases some or all of
them cannot be met. To improve our understanding, then, we will be relaxing
several assumptions (copied from the list at the start of this document), as
follows:
Relaxing assumptions concerning prey-type
characteristics:
w
only one prey type (species) is being attacked
w
all prey individuals are identical/interchangeable (no
age/sex/health/etc. variation)
w
the predator thus takes the same period of
“handling-time” to deal with each prey
We will look at a model of optimal diet-selection or
prey-type choice to relax this set. See pages 225-227 in Krebs.
Relaxing assumptions concerning prey depletion and
predator satiation:
w
the number of prey caught is very small compared to
the number available (i.e. no depletion)
w
the predator is as hungry at the end as it was at the
start (hunts just as hard)
We will develop a graphical model of patch-use
(Marginal Value theory) to relax these. This model approach is not covered
explicitly in Krebs, but adds to our understanding usefully.
Relaxing assumptions concerning number of predators:
w
the model looks at a single individual predator in
isolation
We will develop (mostly graphical)
Lotka-Volterra-type dynamic models of entire predator and prey populations in
interaction with each other. Similar coverage is in Krebs, pages 207-212.