
The Debate over Dominance
"As a rule, hybrids do not represent the form exactly intermediate between the parental strains...Those traits that pass into hybrid association entirely or almost entirely unchanged, thus themselves representing the traits of the hybrid, are termed dominating, and those that become latent in the association, recessive."
- Gregor Mendel (1865)
Mendel's work with peas revealed that certain traits tend to be dominant over others (i.e. expressed whenever inherited), e.g. round pea shape:
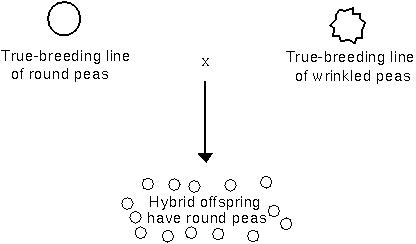
All of the variants examined by Mendel are caused by differences in single genes.
A pea plant carries two copies of each gene (called alleles).
For all seven traits examined by Mendel (pea shape, pea color, flower color, pod shape, pod color, flower position, stem length), one allele was completely dominant (A) and one allele was completely recessive (a).
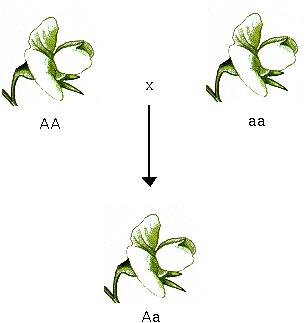
For other traits, individuals with both alleles (heterozygotes, Aa) can be distinguished from both parental forms.
For example, among human conditions: ulcerative colitis, hemolytic anemia, phenylketonuria, familial nonhemolytic jaundice.
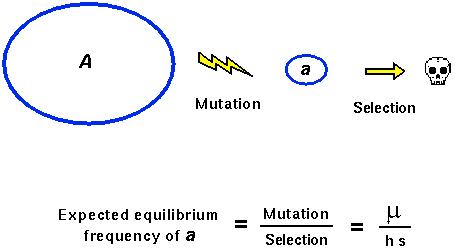
What is the dominance level of new mutant alleles?
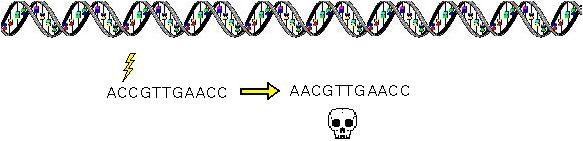
Mutations that occur within the DNA sequence, if they have any effect at all, tend to reduce "fitness".
They also tend to be partially or fully recessive.
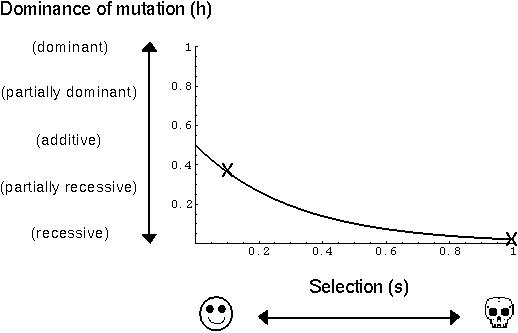
Why do new mutant alleles tend to be recessive while alleles that are common in a population tend to be dominant?
 |
"...there is a tendency always at work in nature which modifies the response of the organism to each mutant gene in such a way that the wild type tends to become dominant."
|

Fisher recognized, however, that there won't generally be many mutant alleles within a population.
Without many mutant alleles, there won't be many heterozygotes and so selection on the wildtype to become more dominant must be weak.
Fisher recognized this problem but argued:
"Nevertheless, considering the ratio of the periods of time available it seems not impossible, but rather probable, that the reaction of the wild type to the heterozygous phase of a recurrent mutation has in some cases at least been modified to an appreciable extent."
- Fisher (1928)
 |
"...can a selection pressure of this order produce any appreciable evolutionary effect, however long it may continue?"
|
"The question at issue reduces then to whether there are genes so neutral in relation to all other evolutionary forces that selections which change their frequencies at rates of the order of mutation pressure are the most important forces acting on them, and whether such genes are sufficiently numerous to give a basis for such a common phenomenon as dominance."
- Wright (1929b)
How did Wright explain the tendency for mutations to be recessive?
"...mutations are most frequently in the direction of inactivation and that for physiological reasons inactivation should generally behave as recessive."
- Wright (1929a)
Wright suggested that many gene products may act as catalysts, and that these would naturally exhibit dominance.
"It is easy to show that increase in the activity of a gene should soon lead to a condition in which even doubling of its immediate effect brings about little or no increase in the ultimate effects."
- Wright (1929a)
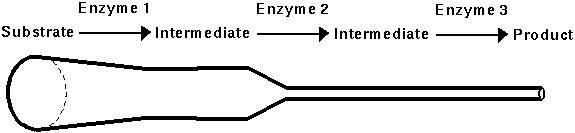
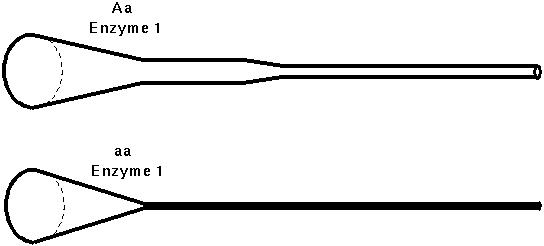
For many enzymes, a small reduction in activity may have little effect, but a large reduction may severely reduce the flow through an enzymatic pathway.
Fisher (1934)
"Professor Sewall Wright, who had perhaps overlooked or misunderstood the calculations in my paper..."
"In a paper published in January, 1934, Wright accepts my correction to his calculation. He does not refer to its history, or to the fact that, in 1928, I had already shown two facts respecting the selection intensities arrived at."
"Professor Wright mentions another argument which should be answered, as it evidently weighs with him, though the fallacy is a simple one. He says "There should always be other evolutionary pressures of greater magnitude acting in one direction or the other," and appears to think that this implies that a selective intensity of lesser magnitude has therefore no effect."
Wright (1934)
"I trust that Dr. Fisher will acknowledge that he also has not always been careful in historical matters."
"Returning to the theory of dominance, Fisher's recent paper (Fisher, 1934) goes on to expose various alleged fallacies in my reasoning. I, in turn, am unable to follow his reasoning but am willing to leave the matter at this point to the reader who wishes to compare his arguments with those which I have expressed..."
"The acrimonious exchange in 1934 between Wright and Fisher over the evolution of dominance marked the end of their previously cordial interchange of ideas. After 1934 they never again corresponded or even had a congenial conversation (stories about their occasional meetings abound), although they did continue to exchange reprints."
Who was right?
Several theoretical and empirical studies have since bolstered Wright's physiological theory of dominance.
Kacser and Burns (1981) developed a model for enzymatic pathways and predicted from it that most enzymes would be dominant.
Models of Fisher's theory suggest that the strength of selection on a modifier of dominance is proportional to  and does not depend on the value of h or s.
and does not depend on the value of h or s.
Models of Wright's theory suggest that mutations of small effect should behave nearly additively, while mutations of large effect will tend to be recessive.
 The observed relationship between dominance and mutant effect supports Wright's theory (Charlesworth 1979)
The observed relationship between dominance and mutant effect supports Wright's theory (Charlesworth 1979)
 |
Chlamydomonas is a normally haploid alga (with only one allele at each gene, e.g. either A or a, but not Aa).
|
"This result falsifies Fisher's theory of dominance and provides strong support for the alternative theory."
Fisher's theory was laid to rest.
Beyond the debate
This debate dichotomized views of the evolution of dominance and led to a surprising lack of investigation with the fertile ground between the views of Fisher and Wright.
"Cases of polymorphism permanently maintained in a species by the stability of the frequency ratio of a pair of allelomorphs supply opportunities peculiarly favourable to the selective evolution of dominance, for in these cases the heterozygotes are not extremely rare."
- Fisher (1931) "If for any reason the proportion of heterozygous mutants reaches the same order as that of the type, selection of modifiers of dominance ...might well become of evolutionary importance."
- Wright (1929a)
Cases where heterozygotes are frequent
Denis Bourguet and I recently examined the last two cases where heterozygotes can be maintained for prolonged periods of time.
Heterozygote Advantage
Imagine that the fitness of different genotypes within a population are measured and divided by the average fitness of the whole population.
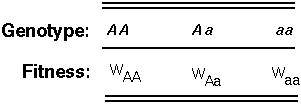
If WAa is greater than both WAA and Waa, the A allele will be maintained by selection within the population at frequency

Heterozygotes (Aa) then occur at frequency 2p(1-p).
We focused on evolutionary changes at a modifier locus (M), whose genotypes affect the fitness of heterozygotes.
A modifier allele, m, that increases WAa by  WAa increases in frequency as if it had a direct selective advantage of:
WAa increases in frequency as if it had a direct selective advantage of:
*
WAa [the extent to which heterozygous fitness is modified]
Selection can strongly favor an allele m that increases fitness within Aa individuals by WAa, even if m causes no direct advantage.
| [If the M and A genes are very close to one another on a chromosome, selection for m goes up by Max[p,1-p]/(2p(1-p)), which =2.3 in this example.] | 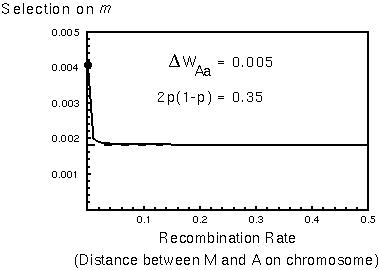 |
Migration-Selection Balance
If the environment favors different types of individuals in different patches, migration between these patches can bring in genetic variation and increase the frequency of heterozygotes (Aa).
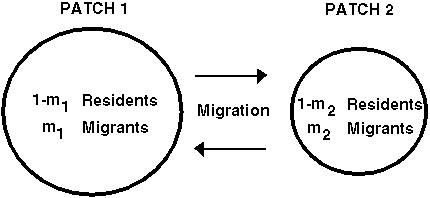
Again we can ask what happens to a modifier allele, m, that affects the level of dominance in one or in both patches:
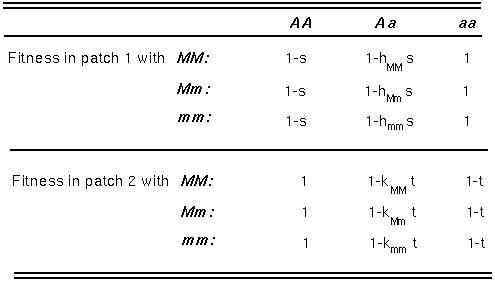
If there is complete mixing of individuals among the patches (high migration), the equations become identical to the case of heterozygote advantage!
Dominance is more likely to evolve:
With lower rates of migration, the analysis is more complex.
Selection on modifiers that alter dominance levels is stronger when heterozygotes are more frequent, but this may occur in species with intermediate levels of migration.
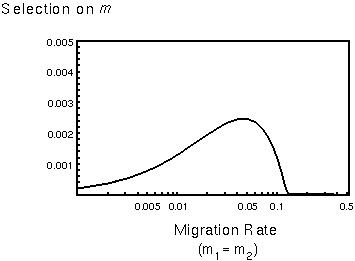
Dominance is as likely to evolve in very productive source patches (those that send out many emigrants and receive few immigrants) as in sink populations (those that send out few emigrants and receive many immigrants).
Selection is always much stronger then at the mutation-selection balance considered by Fisher and could shape the evolution of dominance for any gene that is polymorphic for extended periods of time.
EXAMPLE: The evolution of corn
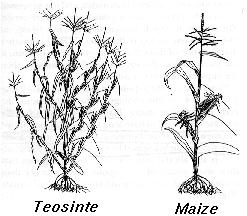
Approximately five major regions of the genome are responsible for the transition from a teosinte-like ancestor to modern corn.
Two of these regions have been identified recently by John Doebley et al. (1995).
One maps to a gene (teosinte branched 1) that affects whether a flower becomes male (tassel) or female (corn cob) and affects the length of side branches.
A second region also affects length of side branches.
By making appropriate crosses between maize and teosinte, the authors were able to place alleles within these regions from maize into a teosinte genetic background.
For every trait that the regions affected, the alleles were less dominant in teosinte than they were in maize.
"This change in gene action could have resulted from selection during the domestication process for modifier loci that enhanced the expression of the trait in the heterozygote."