Lecture 3
Transcription and Chromatin
The Holoenzyme
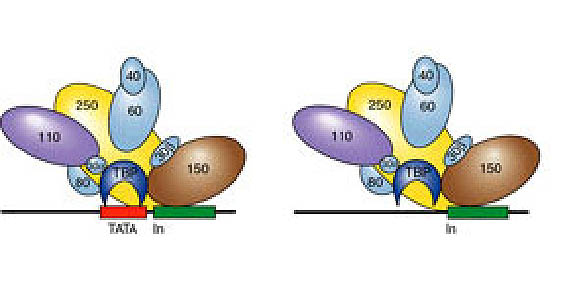
The general transcription factors (GTFs) are TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIH, named in order of discovery. Each of these proteins, with the exception of TFIIB, contains multiple subunits, so the GTFs comprise at least 30 different proteins. TFIID contains the TATA-binding protein (TBP) and a number of TBP-associated proteins (TAFs) generally referred to by their molecular weights (TAFII250, TAFII150 etc.). The TAFs aid in positioning and stabilizing TFIID at particular promoters (TATA-containing and TATA-less), and they alter chromatin structure (TAFII250 is a histone acetyltransferase) at the promoter to allow assembly of GTFs.
Other constituents of the holoenzyme are the Srb proteins. These proteins were originally isolated genetically by a screen for suppressors of RNA PolII mutations. There are multiple Srb proteins and various groups have purified slightly different subsets of these in different holoenzyme preparations. The sequences of Srb proteins have suggested little about their function. Transcription is abolished in yeast mutant for various Srb proteins, showing that they are essential for transcription.
Models for Transcription Initiation
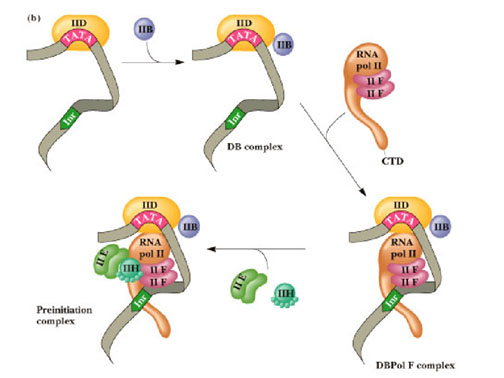
Until recently, basal transcription seemed very straightforward. RNA polymerase II, in association with at least 30 other proteins, were thought to assemble on the TATA box to make a transcriptionally active molecule. More than a decade of studies has left us with a clearer picture of what may be happening. Based on the ability to reconstruct transcriptionally active complexes in vitro, it was determined that an ordered assembly of the transcription pre-initiation complex was required for promoter binding and transcription initiation from naked DNA templates. The steps leading to Pol II transcription are summarized below:
1. TFIID recognizes and binds to TATA promoter element. TFIIB and TFIIA are subsequently recruited.
2. Formation of a more stable complex containing TFIID, TFIIA, TFIIB, hypophosphorylated RNA PolII and TFIIF.
3. An activated complex formed by the addition of TFIIE and TFIIH, which stimulate an ATP-dependent isomerization and promoter-melting event.
4. Promoter clearance and RNA synthesis upon hyperphosphorylation of the RNA PolII CTD (C-terminal domain).
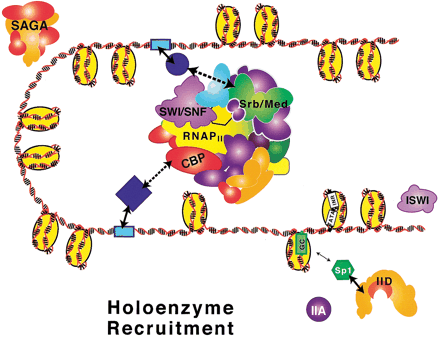
Another popular model, based on the finding that certain preparations of RNA PolII co-purified with subsets co-regulators (discussed later), envisions the targeted recruitment of a completely pre-assembled RNA PolII holoenzyme.
Lemon and Tjian (2000). Genes & Development 14, 2551-2569.
Transcription and Chromatin
Transcriptional activation is mediated by the facilitated binding of the basal transcription complex to the transcription start site of a promoter. The activation procedure involves communication between specific transcription factors and members of the basal transcription complex, perhaps through protein-protein interactions. However, since eukaryotic DNA is packaged into chromatin, the nucleosome being the basic unit, the accessibility of the transcription factors is limited. In order to activate transcription, some of the specific transcription factors must have the capacity to bind to their binding sites within a nucleosomal context. As a next step, the chromatin structure of the promoter needs to be decondensed in order to facilitate the binding of the basal transcription machinery. Recent data have addressed these issues and both binding of transcription factors to their chromatin binding site as well as transcription factor-induced chromatin remodelling have been demonstrated.


The histone tails on the nucleosome surface are subject to enzyme-catalyzed modifications that may, singly or in combination, form a code specifying patterns of gene expression.
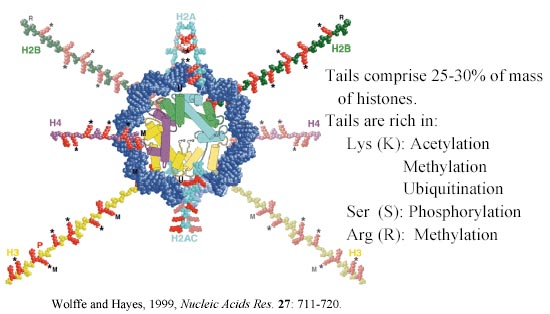

Strahl and Allis (2000). Nature 403, 41-45.
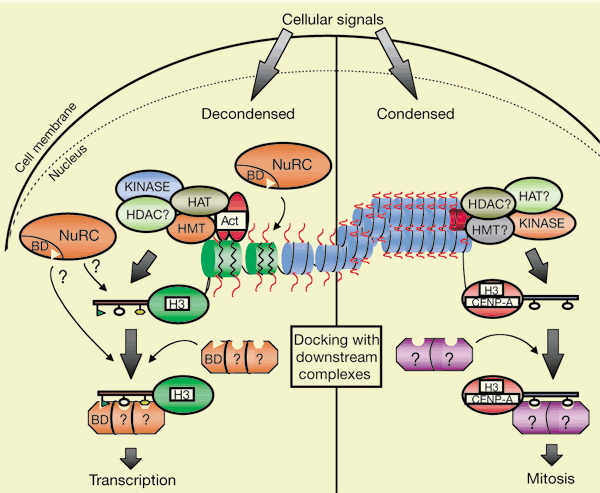
Strahl and Allis (2000). Nature 403, 41-45.
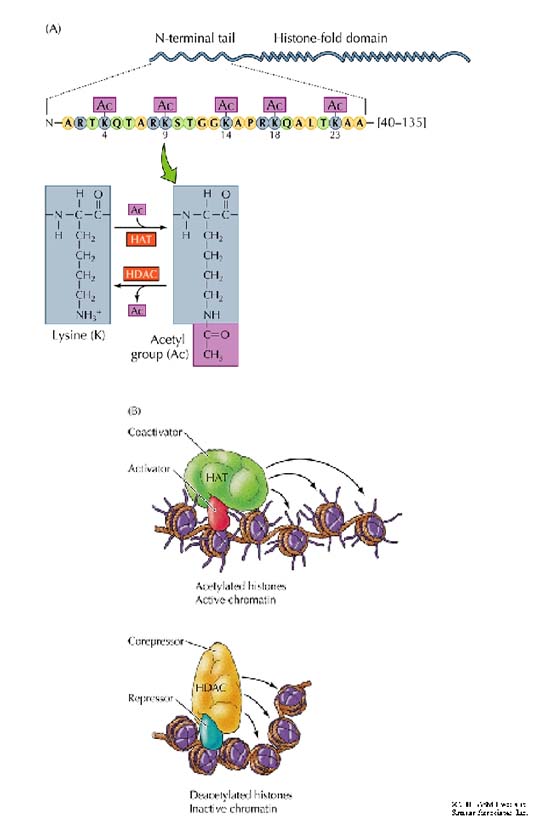
In summary, activators, chromatin-modifying enzymes, and basal transcription factors unite to activate genes. These factors are recruited in a precise order to promoters. The timing of the activation of transcription and the ordered recruitment of factors to promoters are the engines which, at the right moment and for the right length of time, drive the transcriptional regulation of each gene throughout the life of a cell.